|
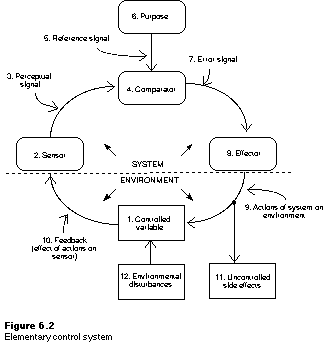
[Above
Figure from Gary Cziko's Home Page]
This page is a compilation
of information about how humans might control their perceptions. Information on
perceptual control theory (PCT) is included to provide a realistic psychological
perspective on neuroscience research. According to perceptual control theory, behavior
is the control of perception rather than the response to a stimulus. Research
involving various types of control systems that can be found in the brain is included
on this page to demonstrate that control systems are not solely in the domain
of engineers. If the concepts of homeostasis and homeorhesis may be applied to
internal environments, can they not also be applied to the external environment
of an organism? The brain is a means toward transferring perceptual signals derived
from the external environment into the internal environment of billions of interconnected
neurons. Control systems within the brain and body are responsible for keeping
perceptual signals within survivable limits, regardless of the nature of the environment
that they are derived from. By integrating
perceptual control theory with research from the field of neuroscience, the future
of human self-awareness might be realized by individuals willing to do
so.
Mansell W.
Control theory and psychopathology: an integrative approach.
Psychol Psychother. 2005 Jun;78(Pt 2):141-78.
Perceptual control theory (PCT; Powers, 1973) is presented and adapted as a framework to understand the causes, maintenance, and treatment of psychological disorders. PCT provides dynamic, working models based on the principle that goal-directed activity arises from a hierarchy of negative feedback loops that control perception through control of the environment. The theory proposes that psychological distress arises from the unresolved conflict between goals. The present paper integrates PCT, control theory, and self-regulatory approaches to psychopathology and psychotherapy and recent empirical findings, particularly in the field of cognitive therapy. The approach aims to offer fresh insights into the role of goal conflict, automatic processes, imagery, perceptual distortion, and loss of control in psychological disorders. Implications for psychological therapy are discussed, including an integration of the existing work on the assessment of control profiles and the use of assertive versus yielding modes of control. [Abstract]
Vancouver JB.
The depth of history and explanation as benefit and bane for psychological control theories.
J Appl Psychol. 2005 Jan;90(1):38-52.
A longstanding debate has recently re-erupted in the self-regulation literature
around the concept of self-efficacy. This article presents an argument that the
debate emerges from a lack of understanding of the history of control theories
within both the social and physical sciences and the various levels of
explanation to which phenomena can be subjected. This history, coupled with the
issues of determinism, materialism, and empiricism evoked by the deeper level of
explanation that some versions of control theory provide, has led some critics
to mis-apply non-psychological properties to control theories and obscure their
usefulness. Here, the usefulness of a deeper control theory level of explanation
is illustrated using comparisons with explanations found in goal-setting theory
and social cognitive theory. [Abstract] [Full Text]
Marken RS.
Controlled
variables: psychology as the center fielder views it.
Am
J Psychol 2001 Summer;114(2):259-81
"Perceptual
control theory (PCT) views behavior as the control of perception. The central
explanatory concept in PCT is the controlled variable, which is a perceived aspect
of the environment that is brought to and maintained in states specified by the
organism. According to PCT, understanding behavior is a matter of discovering
the variables that organisms control. But the possible existence of controlled
variables has been largely ignored in the behavioral sciences. One notable exception
occurs in the study of how baseball outfielders catch fly balls. In these studies
it is taken for granted that the fielder gets to the ball by controlling some
visual aspect of the ball's movement. This article describes the concept of a
controlled variable in the context of research on fly ball catching behavior and
shows how this concept can contribute to our understanding of behavior in general."
[Abstract]
Richard
S. Marken
Looking at behavior through control theory glasses
Review of General Psychology, 6(3):260-270 2002
"Behavior is always seen through
the theoretical preferences of the observer. These preferences act like different
prescriptions for glasses. The most popular glasses use the causal theory prescription,
through which an organism's behavior appears to be the result of external or internal
causes. This article describes glasses that use the less familiar control theory
prescription, through which behavior looks like the organism's purposeful efforts
to control its own perceptions. The consequences of looking at the same behavior
through these different "glasses" are demonstrated by comparing examples
of real-life behavior with the behavior of computer simulations available on the
Internet. A method is described that makes it possible to determine which "glasses"
provide the best view of any particular example of behavior."
Richard
S. Marken
Fielder’s Choice: A Unified Theory of Catching Fly
Balls
"A closed loop exists when the way a system
acts influences what it perceives while what it perceives is influencing the way
it acts. The fielder’s path choice occurs in a closed loop because the way
fielders act (the path chosen) influences what they perceive (the fly ball) while
what they perceive is influencing the way they act. When behavior occurs in a
closed loop, the behaving system is acting as a control system and the appropriate
theoretical framework for understanding the system’s behavior is control
theory. Control theory shows that the behavior of a control system (such as
a fielder) must be understood as a process of controlling perception rather
than choosing action (Powers, 1973).
A control system
acts to bring a perception of some aspect of its environment to a predetermined
or reference state while protecting it from the effects of disturbance.
This process is called control and the perception that is brought to and
maintained in the reference state is called a controlled variable (Powers,
1978). The actions that protect the controlled variable from disturbance are driven
by error -- the difference between the reference and actual state of the controlled
variable -- not by information about the disturbance itself. So the actions of
a control system depend on (often invisible) disturbances to the controlled variable,
not on information regarding the actions to be taken to deal with those disturbances.
From a control theory perspective, therefore, a unified fielder theory must account
for the perceptions a fielder chooses to control rather than the path the fielder
chooses to take." [Full
Text PDF]
Control
Systems Group
Responsible
Thinking Process
(Includes several good articles about PCT) PERCEPTUAL
CONTROL THEORY (PCT):
Basic Models (Includes a nice control system diagram) Bruce
Abbott's PCT Site Gary
Cziko's Home Page
Gary Cziko
Without Miracles: Universal Selection Theory and the Second Darwinian
Revolution
"But of most importance for our purposes,
the theory provides a plausible explanation for how behavior can become and remain
adaptedly complex. It is clearly not the case, as believed by Darwin and Lorenz,
that organisms with useful fixed behaviors are selected during the course of evolution,
resulting in innate, fixed patterns of behavior known as instincts. And it is
also not the case that specific behaviors are selected by the environment by contingencies
of reward during the life of the organism, as believed by Thorndike and Skinner.
It is the selection of organisms with useful, adapted perceptual control systems
over the course of evolution, coupled with the organism's cumulative variation
and election of its own perceptual control systems during its relatively brief
life, that accounts for the adapted nature of behavior. There is no instruction
by the environment, no stamping in of stimulus-response connections within the
nervous system. Rather, we find a very Darwinian process of selection, not of
behaviors, but of closed, negative-feedback loops encompassing perception, comparison
with the reference level, and action, which allow patterns of behavior to remain
functional, not only from one occasion to the next, but also within the continually
changing environment of the behavior itself.
What
may seem mysteriously ironic in all this is to realize that the purposeless process
of natural selection has led to the evolution of purposeful organisms. But the
irony fades when one considers the great survival and reproductive advantages
of organisms that are able consistently to achieve goals essential to their survival
and reproduction despite an unpredictable, uncaring, and often hostile environment."
[Book Link]
Gary Cziko
The Things We Do Using the Lessons of Bernard and
Darwin to Understand the What, How, and Why of Our Behavior
"The basic Darwinian lesson informs us that our evolutionary past provided
us and all animals with certain basic preferences. We prefer certain
foods,
odors, and tastes and are repulsed by others. We prefer environments
that
are not too hot and not too cold. We look for certain characteristics
in mates,
which differ depending on our sex. We do what we can
to assist the well-being
of our children, close relatives, and other individuals
from whom we can expect
such assistance in return. We prefer the
company of family members and others
who are most like us, and are wary
of others whom we perceive as physically,
racially, or culturally different.
But these preferences, naturally selected
for their past survival and reproductive consequences, are not necessarily advantageous
in these respects
in the modern environment we inhabit.
The extended
Bernardian lesson provides an explanation for how such
preferences, existing
as reference levels within feedback-control systems,
influence our behavior,
and how we are able to purposefully vary our
behavior to make our perceptions
match these reference levels." [Book
Link] Vancouver JB, Putka DJ.
Analyzing
Goal-Striving Processes and a Test of the Generalizability of Perceptual Control
Theory.
Organ Behav Hum Decis Process 2000 Jul;82(2):334-362
"Theories that articulate dynamic processes are relatively rare, but methods
for testing the theories are even rarer. This study illustrates two methods for
examining goal-striving processes and a tool for collecting dynamic data. The
first method tests a hypothesis regarding what variable the participants are attempting
to maintain. The second method involves creating multilevel models used to describe
the dynamic data generated by study participants, which can be used to test between-
and within-subject manipulations or differences. The tool is a research simulation
of a manager's role in scheduling subordinates in a hospital wing. Together these
methods and the tool are used to test the generalizability of perceptual control
theory in explaining striving for cognitive goals. The results confirm the viability
of a control theory accounting of goal striving and highlight the potential of
the methods and the research tool in future research." [Abstract]
Kent McClelland
The collective control of perceptions: constructing order from conflict
International Journal of Human-Computer Studies 2004 Jan;64(1): 65-99.
This article offers a new perspective on sociological theory, based on psychological insights from Perceptual Control Theory. After describing this cybernetic model of goal-directed behavior and reviewing its empirical support, I present results from computer simulations applying the model to the social interactions of elementary control agents. My key finding is that agents controlling their own perceptions of a single environmental variable can stabilize it even when their intentions conflict. The concluding section discusses implications of this model of collective control processes for the sources of order, conflict, continuity, and change in social life. [Abstract]
KENT McCLELLAND
PERCEPTUAL CONTROL
AND SOCIAL POWER
Sociological Perspectives, Vol. 37, No.
4, pp. 461-496, 1994.
"This paper explores a new psychological perspective
on human behavior, a cybernetic approach called "perceptual control theory" (PCT). After detailing the PCT model, I demonstrate one application of PCT to
sociological theory by applying this perspective to questions of power and interpersonal
control. I argue that social power should be distinguished from interpersonal
use of force, coercion, incentives, or influence. Rather, power derives from an
alignment of goals by humans acting as independent control systems. The paper
closes with a discussion of connections between PCT and several strands of current
sociological theory." [Full
Text]
David M. Goldstein
Perceptual
Control Theory Psychotherapy
[Article
Link]
W. Thomas Bourbon
Perceptual
Control Theory, Reinforcement Theory,
Countercontrol, and the Responsible
Thinking Process ®
[Article
Link]
W. Thomas Bourbon
Perceptual Control Theory,
Reality Therapy, and the Responsible Thinking Process
[Article
Link] Bruce B. Abbott
A Synopsis
of William T. Powers' Perceptual Control Theory
"HPCT
[Hierarchical Perceptual Control Theory] proposes a second control mechanism lying
outside of the perceptual control hierarchy. Called the reorganizing system, this
mechanism functions to change the organization of the perceptual control hierarchy
so as to remove damaging conflicts and improve control of perceptual signals at
all levels. Reorganization can involve merely changing the parameters of control
(e.g., the loop gain) within a given system so as to increase its efficiency or
sensitivity or eliminate problems such as a tendency to oscillate. Alternatively,
reorganization can involve establishing new connections between levels of the
control hierarchy, destroying existing connections, or even creating new perceptual
signals (by combining next-down perceptual signals in new ways) and new control
systems to control them. If a reorganizing system
is to be effective, it must have a mechanism to determine when changes are needed,
and some means of implementing the changes. Powers proposes (after Ashby, 1952)
that what drives reorganization is persistent error in intrinsic variables. This
presupposes some mechanism capable of sensing the states of these intrinsic variables,
perhaps through chemical means if not via neural signals. According to Powers,
these states are compared to genetically given reference values to compute the
level of error. In Powers' proposal, reorganization occurs continually at a rate
proportional to the level of persistent error. Thus reorganization will proceed
more swiftly when error is large then when error is small, and may nearly cease
entirely when persistent error is kept very low by the perceptual control hierarchy."
[Article Link] Fay
D, Sonnentag S.
Rethinking the effects of stressors: a longitudinal
study on personal initiative.
J Occup Health Psychol 2002
Jul;7(3):221-34
"This study examined the relationship between stressors
at work and personal initiative (PI), one proactive concept of extra-role performance.
Using a control theory framework to describe the stress process, the authors hypothesized
that stressors should be positively related to PI. This departs from findings
of negative relationships between stressors and other types of performance. Furthermore,
curvilinear relationships were tested. The analyses, based on 4 measurement waves
of a longitudinal field study with 172 to 193 participants, showed that stressors
were positively related to subsequent changes in PI; there was no support for
a curvilinear relationship." [Abstract] Karel
Pacák, and Miklós Palkovits
Stressor Specificity of
Central Neuroendocrine Responses: Implications for Stress-Related Disorders
Endocr.
Rev. 22: 502-548
"All of these data suggest the existence of stressor-specific
central pathways that participate differentially in the regulation of sympathoneuronal
and adrenomedullary outflow as well as activity of the HPA axis. Our conclusions
further support our view and suggestions by others (483) that the neural representations
of stress and other disease-related symptoms and signs cannot be described as
a matter of altered function of one brain region or neurotransmitter. Rather,
it reflects the activation of several circuits to orchestrate an optimal pattern
of neuroendocrine and other responses. It is very likely that one of these circuits
is the primary circuit, others are subordinate, and all combine to form the functional
circuit to guarantee maximal plasticity of stress responses during acute as well
as chronic stress conditions. Stressor-specific formation of new connections and
disconnections of subordinate circuits with the primary one, anatomical and biochemical
switches within the functional circuit, previous experience of an organism to
an individual stressor, and genetically programmed neuronal and cellular functions
precisely tune responses to stress and maximally protect from its deleterious
effects. For example, in the response to an acute stressor, the failure of the
primary circuit to function properly may have disastrous effects for an organism
as is evident from neuroendocrine responses in patients with hypothalamic lesions;
failure of subordinate circuits may contribute to milder symptoms and signs."
[Full Text] Buonomano
DV, Merzenich MM.
Cortical plasticity: from synapses to maps.
Annu
Rev Neurosci. 1998;21:149-86.
"It has been clear for almost two decades
that cortical representations in adult animals are not fixed entities, but rather,
are dynamic and are continuously modified by experience. The cortex can preferentially
allocate area to represent the particular peripheral input sources that are proportionally
most used. Alterations in cortical representations appear to underlie learning
tasks dependent on the use of the behaviorally important peripheral inputs that
they represent. The rules governing this cortical representational plasticity
following manipulations of inputs, including learning, are increasingly well understood.
In parallel with developments in the field of cortical map plasticity, studies
of synaptic plasticity have characterized specific elementary forms of plasticity,
including associative long-term potentiation and long-term depression of excitatory
postsynaptic potentials. Investigators have made many important strides toward
understanding the molecular underpinnings of these fundamental plasticity processes
and toward defining the learning rules that govern their induction. The fields
of cortical synaptic plasticity and cortical map plasticity have been implicitly
linked by the hypothesis that synaptic plasticity underlies cortical map reorganization.
Recent experimental and theoretical work has provided increasingly stronger support
for this hypothesis. The goal of the current paper is to review the fields of
both synaptic and cortical map plasticity with an emphasis on the work that attempts
to unite both fields. A second objective is to highlight the gaps in our understanding
of synaptic and cellular mechanisms underlying cortical representational plasticity."
...
"If
there is a preferred flow of information vertically through the cortex, one
would
expect to find different response characteristics in different cortical layers,
since each stage of cortical processing is presumably contributing to the processing
of information and thus transforming the neuronal response characteristics in
some manner. Indeed, in general, receptive fields tend to be larger and responses
tend to be more complex outside of L-IV. Experiments in rat and monkey somatosensory
cortex indicate that the smallest receptive fields are found in L-IV, while supragranular
layers exhibit larger receptive fields than those observed in L-IV, and infragranular
layers exhibit the largest receptive fields (Simons 1978, Chapin 1986, Armstrong-James
& Fox 1987) or sizes equivalent to those in the supragranular layers (Sur
et al 1985). Laminar analysis in cat visual cortex indicates similar patterns.
Gilbert (1977) reported that the smallest receptive fields were in L-IV, intermediate
size fields were inL-III, and the largest receptive fields were in the infragranular
layers. Other interlaminar differences include the observation that the degree
of orientation tuning is also sharper in the supra- and infragranular layers (Chapman&Stryker
1993) and that the proportion of simple cells in the visual cortex is highest
in L-IV, whereas complex cells are found mostly in supra- and infragranular layers
(Hubel & Wiesel 1968, Gilbert 1977). Together these data support the notion
that at each level of cortical processing, the neurons are sampling from a larger
input space, receiving convergent information from the previous level, diverging
out to the next level, and in the process, forming larger and more complexly integrated
and combinatorial receptive fields. In addition to
the vertical flow of information, there is substantial horizontal interconnectivity,
which integrates information from neighboring regions and from specific, more
distant cortical zones (Lorente de N´o 1938). Excitatory horizontal projections
arise mainly from L-II/III and L-V pyramidal cells and project preferentially
to supra- and infragranular layers (e.g. Schwark & Jones 1989, White 1989,
Abeles 1991, Tanifuji et al 1994). For any given layer, it is not clear what percentage
of synapses originate from within the same cortical column, from more distant
cortical regions, or from other cortical fields. However, even in L-IV, only 15–20%
of the synapses are of thalamic origin (LeVay & Gilbert 1976, Benshalom &
White 1986); most synapses seem to originate from intra- and interlaminar neurons.
Horizontal connectivity may be of particular relevance in cortical map reorganization,
since it appears that areas that develop novel receptive fields and other emergent
response properties after peripheral input manipulations may rely in large part
on connections from neighboring cortical sectors..." [Abstract]
[PDF]
[Abstract]
[PDF]
Biology Online: Regulation
in Biological Systems Hahnloser RH, Douglas RJ,
Hepp K.
Attentional recruitment of inter-areal recurrent networks
for selective gain control.
Neural Comput 2002 Jul;14(7):1669-89
"There is strong anatomical and physiological evidence that neurons with
large receptive fields located in higher visual areas are recurrently connected
to neurons with smaller receptive fields in lower areas. We have previously described
a minimal neuronal network architecture in which top-down attentional signals
to large receptive field neurons can bias and selectively read out the bottom-up
sensory information to small receptive field neurons (Hahnloser, Douglas, Mahowald,
& Hepp, 1999). Here we study an enhanced model, where the role of attention
is to recruit specific inter-areal feedback loops (e.g., drive neurons above firing
threshold). We first illustrate the operation of recruitment on a simple example
of visual stimulus selection. In the subsequent analysis, we find that attentional
recruitment operates by dynamical modulation of signal amplification and response
multistability. In particular, we find that attentional stimulus selection necessitates
increased recruitment when the stimulus to be selected is of small contrast and
of small distance away from distractor stimuli. The selectability of a low-contrast
stimulus is dependent on the gain of attentional effects; for example, low-contrast
stimuli can be selected only when attention enhances neural responses. However,
the dependence of attentional selection on stimulus-distractor distance is not
contingent on whether attention enhances or suppresses responses. The computational
implications of attentional recruitment are that cortical circuits can behave
as winner-take-all mechanisms of variable strength and can achieve close to optimal
signal discrimination in the presence of external noise." [Abstract]
[PDF]
Burrone J, Murthy VN.
Synaptic gain
control and homeostasis.
Curr Opin Neurobiol. 2003 Oct;13(5):560-7.
"Chronic
changes in activity can induce neurons to alter the strength of all their synapses
in unison. Although the specific changes that occur appear to vary depending on
the experimental preparation, their net effect is to counter the experimentally
induced modification of activity. Such adaptive, cell-wide changes in synaptic
strength serve to stabilize neuronal activity and are collectively referred to
as homeostatic synaptic plasticity. Recent studies have shed light on what triggers
homeostatic synaptic plasticity, whether or not it is distinct from other forms
of synaptic plasticity and whether or not it occurs in the intact brain."
[Abstract]
[PDF] Davis
GW, Bezprozvanny I.
Maintaining the stability of neural function:
a homeostatic hypothesis.
Annu Rev Physiol. 2001;63:847-69.
"The
precise regulation of neural excitability is essential for proper nerve cell,
neural circuit, and nervous system function. During postembryonic development
and throughout life, neurons are challenged with perturbations that can alter
excitability, including changes in cell size, innervation, and synaptic input.
Numerous experiments demonstrate that neurons are able to compensate for these
types of perturbation and maintain appropriate levels of excitation. The mechanisms
of compensation are diverse, including regulated changes to synaptic size, synaptic
strength, and ion channel function in the plasma membrane. These data are evidence
for homeostatic regulatory systems that control neural excitability. A model of
neural homeostasis suggests that information about cell activity, cell size, and
innervation is fed into a system of cellular monitors. Intracellular- and intercellular-signaling
systems transduce this information into regulated changes in synaptic and ion
channel function. This review discusses evidence for such a model of homeostatic
regulation in the nervous system." [Abstract] Abbott
LF, Varela JA, Sen K, Nelson SB.
Synaptic depression and cortical
gain control.
Science. 1997 Jan 10;275(5297):220-4.
"Cortical
neurons receive synaptic inputs from thousands of afferents that fire action potentials
at rates ranging from less than 1 hertz to more than 200 hertz. Both the number
of afferents and their large dynamic range can mask changes in the spatial and
temporal pattern of synaptic activity, limiting the ability of a cortical neuron
to respond to its inputs. Modeling work based on experimental measurements indicates
that short-term depression of intracortical synapses provides a dynamic gain-control
mechanism that allows equal percentage rate changes on rapidly and slowly firing
afferents to produce equal postsynaptic responses. Unlike inhibitory and adaptive
mechanisms that reduce responsiveness to all inputs, synaptic depression is input-specific,
leading to a dramatic increase in the sensitivity of a neuron to subtle changes
in the firing patterns of its afferents." [Abstract] Koch
C, Segev I.
The role of single neurons in information processing.
Nat
Neurosci. 2000 Nov;3 Suppl:1171-7.
"Neurons carry out the many operations
that extract meaningful information from sensory receptor arrays at the organism's
periphery and translate these into action, imagery and memory. Within today's
dominant computational paradigm, these operations, involving synapses, membrane
ionic channels and changes in membrane potential, are thought of as steps in an
algorithm or as computations. The role of neurons in these computations has evolved
conceptually from that of a simple integrator of synaptic inputs until a threshold
is reached and an output pulse is initiated, to a much more sophisticated processor
with mixed analog-digital logic and highly adaptive synaptic elements." [Abstract]
[PDF] Rainer
G, Asaad WF, Miller EK.
Selective representation of relevant information
by neurons in the primate prefrontal cortex.
Nature 1998
Jun 11;393(6685):577-9
"The severe limitation of the capacity of working
memory, the ability to store temporarily and manipulate information, necessitates
mechanisms that restrict access to it. Here we report tests to discover whether
the activity of neurons in the prefrontal (PF) cortex, the putative neural correlate
of working memory, might reflect these mechanisms and preferentially represent
behaviourally relevant information. Monkeys performed a 'delayed-matching-to-sample'
task with an array of three objects. Only one of the objects in the array was
relevant for task performance and the monkeys needed to find that object (the
target) and remember its location. For many PF neurons, activity to physically
identical arrays varied with the target location; the location of the non-target
objects had little or no influence on activity. Information about the target location
was present in activity as early as 140ms after array onset. Also, information
about which object was the target was reflected in the sustained activity of many
PF neurons. These results suggest that the prefrontal cortex is involved in selecting
and maintaining behaviourally relevant information." [Abstract] Mao
A, Freeman KA, Tallarida RJ.
Transient loss of dopamine autoreceptor
control in the presence of highly potent dopamine agonists.
Life Sci 1996;59(21):PL317-24
"The concentrations of endogenous ligands
generally remain in a bounded range around a basal level, a manifestation of control.
The dopaminergic system is an excellent example of a control system in which a
negative feedback signal is associated with receptor occupancy of a D2-like dopamine
autoreceptor. A consequence of the control theory is that autoreceptor occupancy
by an agonist results in dopamine levels below the basal, whereas similar stimulation
by a dopamine competitive antagonist results in an increase of dopamine to levels
above the basal. These consequences of control theory were tested and verified
in the rat striatum by infusing graded doses of either the agonist, quinpirole,
or the antagonist, sulpiride, into the rat striatum via a microdialysis probe
and sampling dopamine and metabolite levels at various times after the start of
infusion. Control was maintained even at the very highest doses of these compounds,
i.e., striatal dopamine concentration rose in response to the antagonist and fell
in response to the agonist. In contrast, administration of each of two high affinity
dopamine agonists, 7-OH-DPAT and PPHT showed dose-dependent control only up to
certain doses. Above these doses the dopamine concentration actually increased
to levels well above basal, an indication of loss of control. These findings suggest
that the control of this endogenous ligand does not extend to the very highest
levels of autoreceptor occupancy." [Abstract] Durstewitz,
Daniel, Kelc, Marian, Gunturkun, Onur
A Neurocomputational Theory
of the Dopaminergic Modulation of Working Memory Functions
J. Neurosci. 1999 19: 2807-2822
"The dopaminergic modulation of neural
activity in the prefrontal cortex (PFC) is essential for working memory. Delay-activity
in the PFC in working memory tasks persists even if interfering stimuli intervene
between the presentation of the sample and the target stimulus. Here, the hypothesis
is put forward that the functional role of dopamine in working memory processing
is to stabilize active neural representations in the PFC network and thereby to
protect goal-related delay-activity against interfering stimuli. To test this
hypothesis, we examined the reported dopamine-induced changes in several biophysical
properties of PFC neurons to determine whether they could fulfill this function.
An attractor network model consisting of model neurons was devised in which the
empirically observed effects of dopamine on synaptic and voltage-gated membrane
conductances could be represented in a biophysically realistic manner. In the
model, the dopamine-induced enhancement of the persistent Na+ and reduction of
the slowly inactivating K+ current increased firing of the delay-active neurons,
thereby increasing inhibitory feedback and thus reducing activity of the "background"
neurons. Furthermore, the dopamine-induced reduction of EPSP sizes and a dendritic
Ca2+ current diminished the impact of intervening stimuli on current network activity.
In this manner, dopaminergic effects indeed acted to stabilize current delay-activity.
Working memory deficits observed after supranormal D1-receptor stimulation could
also be explained within this framework. Thus, the model offers a mechanistic
explanation for the behavioral deficits observed after blockade or after supranormal
stimulation of dopamine receptors in the PFC and, in addition, makes some specific
empirical predictions." [Full
Text] Le Bars, Daniel, Gozariu, Manuela, Cadden,
Samuel W.
Animal Models of Nociception
Pharmacol
Rev 2001 53: 597-652
"Sensory systems have the role of informing the
brain about the state of the external environment and the internal milieu of the
organism. Pain is a perception, and as such, it is one of the outputs of a system
in more highly evolved animals -- the nociceptive system -- which itself is a
component of the overall set of controls responsible for homeostasis." [Full
Text] Melzack R.
From the gate to
the neuromatrix.
Pain 1999 Aug;Suppl 6:S121-6
"The
gate control theory's most important contribution to understanding pain was its
emphasis on central neural mechanisms. The theory forced the medical and biological
sciences to accept the brain as an active system that filters, selects and modulates
inputs. The dorsal horns, too, were not merely passive transmission stations but
sites at which dynamic activities (inhibition, excitation and modulation) occurred.
The great challenge ahead of us is to understand brain function. I have therefore
proposed that the brain possesses a neural network--the body-self neuromatrix--which
integrates multiple inputs to produce the output pattern that evokes pain. The
body-self neuromatrix comprises a widely distributed neural network that includes
parallel somatosensory, limbic and thalamocortical components that subserve the
sensory-discriminative. affective-motivational and evaluative-cognitive dimensions
of pain experience. The synaptic architecture of the neuromatrix is determined
by genetic and sensory influences. The 'neurosignature' output of the neuromatrix--patterns
of nerve impulses of varying temporal and spatial dimensions--is produced by neural
programs genetically build into the neuromatrix and determines the particular
qualities and other properties of the pain experience and behavior. Multiple inputs
that act on the neuromatrix programs and contribute to the output neurosignature
include. (1) sensory inputs (cutaneous, visceral and other somatic receptors);
(2) visual and other sensory inputs that influence the cognitive interpretation
of the situation; (3) phasic and tonic cognitive and emotional inputs from other
areas of the brain; (4) intrinsic neural inhibitory modulation inherent in all
brain function; (5) the activity of the body's stress-regulation systems, including
cytokines as well as the endocrine, autonomic, immune and opioid systems. We have
traveled a long way from the psychophysical concept that seeks a simple one-to-one
relationship between injury and pain. We now have a theoretical framework in which
a genetically determined template for the body-self is modulated by the powerful
stress system and the cognitive functions of the brain, in addition to the traditional
sensory inputs." [Abstract] Reichling
DB, Levine JD.
The primary afferent nociceptor as pattern generator.
Pain 1999 Aug;Suppl 6:S103-9
"One of the most important advances in our
understanding of the pain experience was the introduction of the 'gate control'
theory which stimulated analysis of activity pattern in nociceptive pathways and
its modulation. Advances in cellular and molecular biology have recently begun
to provide detailed information on the mechanisms of stimulus transduction within
primary afferent nociceptors as well as mechanisms that modulate the transduction
process. From these new insights into the sensory physiology of the nociceptive
nerve ending emerges a concept of the primary afferent as the first site of pattern
generation in the nociceptive pathway, in which dynamic tuning of gain in the
mosaic of inputs to individual primary afferents occurs. The electrical properties
of the nociceptor membrane that converts the generator potential to a pattern
of action potentials is also actively adjusted." [Abstract] Kobayashi
S.
Warm- and cold-sensitive neurons inactive at normal core temperature
in rat hypothalamic slices.
Brain Res 1986 Jan 1;362(1):132-9
"Electrical activities of thermosensitive neurons were recorded extracellularly
in slices of rat preoptic area and anterior hypothalamus. Of 63 spontaneously
firing neurons found at high searching temperature (37-40 degrees C), 33% were
warm-sensitive, 8% were cold-sensitive and the remaining 59% were thermally insensitive.
In particular, 6 warm-sensitive neurons were active only above 38 degrees C of
rat normal core temperature. In contrast, of 38 spontaneously firing neurons found
at low searching temperature (32-36 degrees C), 8% were warm-sensitive, 29% were
cold-sensitive and the remaining 63% were thermally insensitive. Furthermore,
all these cold-sensitive neurons were active only below 38 degrees C. Therefore,
the warm- and cold-sensitive neurons active at 38 degrees C would be functioning
for narrow band control and the remaining warm- and cold-sensitive neurons inactive
at 38 degrees C would be recruited for wide band control when core temperature
was changed critically from 38 degrees C. Their firing rate activities often showed
obvious threshold responses, large hysteresis of the threshold responses and remarkable
transient responses to slice temperature changes. From aspects of automatic control
theory, these warm- and cold-sensitive neurons themselves may be thermostats to
regulate the brain temperature rather than thermosensors to monitor it."
[Abstract] Sun
JR, Ma YC, Xu ZH, Zhao WJ, Cai YP.
[Effect of norepinephrine on
the thermosensitive neurons in preoptic area of hypothalamus tissue slices in
cold acclimatized rats]
Sheng Li Xue Bao 1997 Dec;49(6):666-70
"In this work, single unit firing activities were recorded in the preoptic
anterior hypothalamus (POAH) brain slices of cold acclimatized and room-temperature
housed rats (CR and RR) and the effects of NE on the neuronal discharges were
observed. The neurons of POAH in CR became much more sensitive to NE than that
in RR (the threshold concentration of NE of CR became significantly lowered).
In comparison with RR, the percentage of warm sensitive neurons that could be
excited by NE was decreased and some of them even showed inhibitory responses.
On the other hand, the percentage of cold sensitive neurons that could be inhibited
by NE was decreased and some of them even showed excitatory responses. The percentage
of thermo-insensitive neurons that could be either excited or inhibited by NE
were increased." [Abstract] Meyer-Lindenberg
A, Ziemann U, Hajak G, Cohen L, Berman KF.
Transitions between
dynamical states of differing stability in the human brain.
Proc Natl Acad Sci U S A 2002 Aug 20;99(17):10948-53
"What mechanisms
underlie the flexible formation, adaptation, synchronization, and dissolution
of large-scale neural assemblies from the 10(10) densely interconnected, continuously
active neurons of the human brain? Nonlinear dynamics provides a unifying perspective
on self-organization. It shows that the emergence of patterns in open, nonequilibrium
systems is governed by their stability in response to small disturbances and predicts
macroscopic transitions between patterns of differing stability. Here, we directly
demonstrate that such transitions can be elicited in the human brain by interference
at the neural level. As a probe, we used a classic motor coordination paradigm
exhibiting well described movement states of differing stability. Functional neuroimaging
identified premotor (PMA) and supplementary motor (SMA) cortices as having neural
activity linked to the degree of behavioral instability. These regions then were
transiently disturbed with graded transcranial magnetic stimulation, which caused
sustained and macroscopic behavioral transitions from the less stable out-of-phase
to the stable in-phase movement, whereas the stable pattern could not be affected.
Moreover, the strength of the disturbance needed (a measure of neural stability)
was linked to the degree of behavioral stability, demonstrating the applicability
of nonlinear system theory as a powerful predictor of the dynamical repertoire
of the human brain." [Abstract] Abe
H.
Nutrition and control theory.
J Nutr
Sci Vitaminol (Tokyo) 1991 Dec;37 Suppl:S5-12
"Organ homeostasis in humans
is maintained by the "auto-regulatory system." Although this biological
regulatory system is regarded as complicated and difficult to analyze, with the
aid of control theory it has become possible for us to construct a mathematical
model. To date we have created models for water-electrolyte balance and glucose
metabolism. The validity of these models has been evaluated by simulation study.
As a clinical application of the model, we have successfully developed an artificial
pancreas system for the treatment of diabetic patients. Through this system, we
have made many contributions to the study of clinical nutrition and we feel that
this has initiated a new era in this field. In order to make further progress
in this field, it will be necessary to take a new dynamic and systemic approach."
[Abstract]
Peters A, Schweiger U, Fruhwald-Schultes B, Born J,
Fehm HL.
The neuroendocrine control of glucose allocation.
Exp Clin Endocrinol Diabetes 2002 Jul;110(5):199-211
"Summary. Here we
propose that glucose metabolism can be understood on the basis of three concept-derived
axioms: (I) A hierarchy exists among the glucose-utilizing organs with the brain
served first, followed by muscle and fat. (II) Tissue-specific glucose transporters
allocate glucose among organs in order to maintain brain glucose concentrations.
(III) Exogenous carbohydrate supply compensates for glucose alterations that can
temporarily occur in muscle and fat. Derived from the control theory, the simplest
solution of allocating supply to 2 organs, e.g. brain and muscle, is a "fishbone"-structured
model. We reviewed the literature, searching for neuroendocrine and metabolic
mechanisms that can fulfill control functions in such a model: The tissue-specific
glucose transporters are differentially regulated. GLUT 1, carrying glucose across
the blood-brain-barrier, is independent of insulin. Instead, this trans-endothelial
glucose transporter is rather dependent on potent regulators of blood vessel function
like vascular endothelial growth factor - a pituitary counterregulatory hormone.
GLUT 4, carrying glucose across the membranes of muscle and fat cells, depends
on insulin. Thereby, insulin allocates glucose to muscle and fat. The hypothalamus-pituitary-adrenal
(HPA) axis, the sympathetic nervous system (SNS), and vascular endothelial growth
factor allocate glucose to the brain. Multiple "sensors" (some of which
have only recently been identified as ATP sensitive potassium channels) measure
glucose or glucose equivalents at various sites of the body: the ventromedial
hypothalamus, the lateral hypothalamus, portal vein, pancreatic beta cell, renal
tubule, muscle and adipose tissue. Feedback pathways both from the brain and from
muscle and fat are involved in regulating glucose allocation and exogenous glucose
supply. The main feedback signal from the brain is found to be glucose, that from
muscle and fat appears to be leptin. In fact, the literature search revealed two
or more biological mechanisms for the function of each component in the model,
finding glucose regulation highly redundant. This review focuses on "brain
glucose" control. The concept of glucose allocation presented here challenges
the common opinion of "blood glucose" being the main parameter controlled.
According to the latter opinion, hyperglycemia in the metabolic syndrome is due
to a putative defect located within the closed loop including the beta cell, muscle
and fat cells. That traditional view leaves some peculiarities of e.g. the metabolic
syndrome unexplained. The concept of glucose allocation, however, would predict
that weight gain - with abundance of glucose in muscle and fat - increases feedback
to the brain (via hyperleptinemia) which in turn results in HPA-axis and SNS overdrive,
impaired insulin secretion, and insulin resistance. HPA-axis overdrive would account
for metabolic abnormalities such as central adiposity, hyperglycemia, dyslipidemia,
and hypertension, that are well known clinical aspects the metabolic syndrome.
This novel viewpoint of "brain glucose" control may shed new light on
the pathogenesis of the metabolic syndrome and type 2 diabetes." [Abstract] Heap
RB, Galil AK, Harrison FA, Jenkin G, Perry JS.
Progesterone and
oestrogen in pregnancy and parturition: comparative aspects and hierarchical control.
Ciba Found Symp 1977;(47):127-57
"The different ways in which the progesterone
requirements of pregnancy are met in various species are reviewed here. Progesterone
production expressed in terms of metabolic body weight lies within about one order
of magnitude in several species (but not in the rat), whether the hormone is predominantly
ovarian or placental in origin. Parturition is usually preceded by a decrease
in the plasma concentration of progesterone and the evidence which suggests that
a decrease in secretion involves enzyme induction is summarized. In the sheep
a sequence of well-marked hormonal changes can be discerned--an increase in fetal
cortisol secretion followed by a fall in placental progesterone and a rise in
oestrogen and prostaglandin F2alpha secretion. This sequence has been interpreted
in terms of hierarchical control theory which envisages a series of levels placed
in a descending order of importance and in which there is a downward transmission
of a signal from a higher level that modifies activity at a lower level. A dominant
level in the hierarchy in the sheep and pig seems to be the fetal hypothalamus
and pituitary, but in the rabbit the maternal hypothalamus, pituitary and ovary
play a more dominant role." [Abstract]
Gerald
M. Edelman, and Joseph A. Gally
Degeneracy and complexity in biological
systems
PNAS 98: 13763-13768; published online before print
as 10.1073/pnas.231499798
"Degeneracy, the ability of elements that are
structurally different to perform the same function or yield the same output,
is a well known characteristic of the genetic code and immune systems. Here, we
point out that degeneracy is a ubiquitous biological property and argue that it
is a feature of complexity at genetic, cellular, system, and population levels.
Furthermore, it is both necessary for, and an inevitable outcome of, natural selection."
[Full Text] Giulio
Tononi, Olaf Sporns, and Gerald M. Edelman
Measures of degeneracy
and redundancy in biological networks
PNAS 96: 3257-3262,
1999.
"Degeneracy, the ability of elements that are structurally different
to perform the same function, is a prominent property of many biological systems
ranging from genes to neural networks to evolution itself. Because structurally
different elements may produce different outputs in different contexts, degeneracy
should be distinguished from redundancy, which occurs when the same function is
performed by identical elements. However, because of ambiguities in the distinction
between structure and function and because of the lack of a theoretical treatment,
these two notions often are conflated. By using information theoretical concepts,
we develop here functional measures of the degeneracy and redundancy of a system
with respect to a set of outputs. These measures help to distinguish the concept
of degeneracy from that of redundancy and make it operationally useful. Through
computer simulations of neural systems differing in connectivity, we show that
degeneracy is low both for systems in which each element affects the output independently
and for redundant systems in which many elements can affect the output in a similar
way but do not have independent effects. By contrast, degeneracy is high for systems
in which many different elements can affect the output in a similar way and at
the same time can have independent effects. We demonstrate that networks that
have been selected for degeneracy have high values of complexity, a measure of
the average mutual information between the subsets of a system. These measures
promise to be useful in characterizing and understanding the functional robustness
and adaptability of biological networks." [Full
Text] Emmons RA.
Abstract versus
concrete goals: personal striving level, physical illness, and psychological well-being.
J Pers Soc Psychol 1992 Feb;62(2):292-300
"This study examined the relation
between personal striving level and psychological and physical well-being. Level
refers to the degree of generality versus specificity of one's goal strivings.
In 3 studies, 188 Ss generated lists of their personal strivings, which were then
rated on specificity level. High-level striving was associated with more psychological
distress, particularly depression. Low-level striving was related to higher levels
of physical illness. Correlations between striving level and self-reported symptoms
were generally not as strong as those between level and the more objective illness
indicators. High-level strivings were seen as more difficult and requiring more
effort than low-level strivings. Results are interpreted in terms of control theory,
goal-setting theory, and the repressive personality style." [Abstract] Vancouver
JB, Thompson CM, Tischner EC, Putka DJ.
Two studies examining the
negative effect of self-efficacy on performance.
J Appl
Psychol 2002 Jun;87(3):506-16
"Although hundreds of studies have found
a positive relationship between self-efficacy and performance, several studies
have found a negative relationship when the analysis is done across time (repeated
measures) rather than across individuals. W. T. Powers (1991) predicted this negative
relationship based on perceptual control theory. Here, 2 studies are presented
to (a) confirm the causal role of self-efficacy and (b) substantiate the explanation.
In Study 1, self-efficacy was manipulated for 43 of 87 undergraduates on an analytic
game. The manipulation was negatively related to performance on the next trial.
In Study 2, 104 undergraduates played the analytic game and reported self-efficacy
between each game and confidence in the degree to which they had assessed previous
feedback. As expected, self-efficacy led to overconfidence and hence increased
the likelihood of committing logic errors during the game." [Abstract] Kim
Y.
The role of cognitive control in mediating the effect of stressful
circumstances among Korean immigrants.
Health Soc Work
2002 Feb;27(1):36-46
"The study reported in this article investigated
relationships among stressful circumstances, cognitive control (the individual's
perception of control over life), and distress among Korean immigrants in the
United States. Specifically, it was hypothesized that cognitive control mediated
the effect of exposure to stressful circumstances on distress. A total of 159
Korean immigrants participated in the study. The result of the study provided
strong support for the role of cognitive control in mediating the relationship
between stressful circumstances and distress. The detrimental effect of exposure
to stressful circumstances on distress was weakened by cognitive control."
[Abstract]
Coping with unplanned childhood hospitalization: the mediating functions
of parental beliefs.
J Pediatr Psychol 1995 Jun;20(3):299-312
"Examined the processes by which two types of informational interventions
(child behavioral information and parental role information) exerted effects on
the coping outcomes of 108 mothers whose young children experienced unplanned
hospitalization. Driven by a strong theoretical framework comprising self-regulation
theory and control theory, this study's findings revealed that the effects of
the experimental interventions were mediated by parental beliefs about their children's
likely behavioral changes and their parental role during hospitalization. Thus,
progress was made in beginning to understand how informational interventions actually
enhance parent coping with childhood hospitalization." [Abstract]
Kobak RR, Cole HE, Ferenz-Gillies R, Fleming WS, Gamble
W.
Attachment and emotion regulation during mother-teen problem
solving: a control theory analysis.
Child Dev 1993 Feb;64(1):231-45
"We present a control theory analysis of adolescents' attachment strategies
in the Adult Attachment Interview (AAI). In Study 1, Q-sort prototypes for secure/anxious
and deactivating/hyperactivating strategies were used to differentiate between
Main and Goldwyn's AAI classifications. In Study 2, we examined how AAI strategies
were associated with emotion regulation during mother-teen problem solving. 4
aspects of mother-teen problem solving (dysfunctional anger, support/validation,
avoidance of problem solving, and maternal dominance) were used to predict teens'
AAI strategies. Teens with secure strategies engaged in problem-solving discussions
characterized by less dysfunctional anger and less avoidance of problem solving.
In addition, attachment security showed a curvilinear relation with maternal dominance,
indicating that secure teens maintained balanced assertiveness with their mothers.
Teens with deactivating strategies engaged in problem-solving interactions characterized
by higher levels of maternal dominance and dysfunctional anger. The contribution
of attachment strategies to teens' autonomy and to transformations in mother-teen
relationships is discussed." [Abstract] |
Posner MI, Rothbart MK.
Attention,
self-regulation and consciousness.
Philos Trans R Soc Lond
B Biol Sci. 1998 Nov 29;353(1377):1915-27.
"Consciousness has many aspects.
These include awareness of the world, feelings of control over one's behaviour
and mental state (volition), and the notion of continuing self. Focal (executive)
attention is used to control details of our awareness and is thus closely related
to volition. Experiments suggest an integrated network of neural areas involved
in executive attention. This network is associated with our voluntary ability
to select among competing items, to correct error and to regulate our emotions.
Recent neuroimaging studies suggest that these various functions involve separate
areas of the anterior cingulate. We have adopted a strategy of using marker tasks,
shown to activate the brain area by imaging studies, as a means of tracing the
development of attentional networks. Executive attention appears to develop first
to regulate distress during the first year of life. During later childhood the
ability to regulate conflict among competing stimuli builds upon the earlier cingulate
anatomy to provide a means of cognitive control. During childhood the activation
of cingulate structures relates both to the child's success on laboratory tasks
involving conflict and to parental reports of self-regulation and emotional control.
These studies indicate a start in understanding the anatomy, circuitry and development
of executive attention networks that serve to regulate both cognition and emotion."
[Abstract] Aarts
H, Dijksterhuis A, De Vries P.
On the psychology of drinking: being
thirsty and perceptually ready.
Br J Psychol 2001 Nov;92(Pt
4):631-42
"The present research is concerned with cognitive effects of
habitually regulated primary motives. Specifically, two experiments tested the
idea that feelings of thirst enhance the cognitive accessibility of, or readiness
to perceive, action-relevant stimuli. In a task allegedly designed to assess mouth-detection
skills, some participants were made to feel thirsty, whereas others were not.
Results showed that participants who were made thirsty responded faster to drinking-related
items in a lexical decision task, and performed better on an incidental recall
task of drinking-related items, relative to no-thirst control participants. These
results suggest that basic needs and motives, such as thirst, causes a heightened
perceptual readiness to environmental cues that are instrumental in satisfying
these needs." [Abstract] Toppino
TC.
Reversible-figure perception: mechanisms of intentional control.
Percept
Psychophys. 2003 Nov;65(8):1285-95.
"Observers can exert a degree of intentional
control over the perception of reversible figures. Also, the portion of the stimulus
that is selected for primary or enhanced processing (focal-feature processing)
influences how observers perceive a reversible figure. Two experiments investigated
whether voluntary control over perception of a Necker cube could be explained
in terms of intentionally selecting appropriate focal features within the stimulus
for primary processing. In Experiment 1, varying observers' intentions and the
focus of primary processing produced additive effects on the percentage of time
that one alternative was perceived. In Experiment 2, the effect of varying the
focus of primary processing was eliminated by the use of a small cube, but the
effect of intention was unaltered. The results indicate that intentional control
over perception can be exerted independently of focal-feature processing, perhaps
by top-down activation or priming of perceptual representations. The results also
reveal the limits of intentional control." [Abstract] Gilbert
CD, Sigman M, Crist RE.
The neural basis of perceptual learning.
Neuron 2001 Sep 13;31(5):681-97
"Perceptual learning is a lifelong process.
We begin by encoding information about the basic structure of the natural world
and continue to assimilate information about specific patterns with which we become
familiar. The specificity of the learning suggests that all areas of the cerebral
cortex are plastic and can represent various aspects of learned information. The
neural substrate of perceptual learning relates to the nature of the neural code
itself, including changes in cortical maps, in the temporal characteristics of
neuronal responses, and in modulation of contextual influences. Top-down control
of these representations suggests that learning involves an interaction between
multiple cortical areas." [Abstract] Smith,
Edward E., Jonides, John
Storage and Executive Processes in the
Frontal Lobes
Science 1999 283: 1657-1661
"Most
researchers concur that executive processes are mediated by PFC and are involved
in the regulation of processes operating on the contents of working memory. Although
there is lack of consensus about a taxonomy of executive processes, there is some
agreement that they include (i) focusing attention on relevant information and
processes and inhibiting irrelevant ones ("attention and inhibition");
(ii) scheduling processes in complex tasks, which requires the switching of focused
attention between tasks ("task management"); (iii) planning a sequence
of subtasks to accomplish some goal ("planning"); (iv) updating and
checking the contents of working memory to determine the next step in a sequential
task ("monitoring"); and (v) coding representations in working memory
for time and place of appearance ("coding")." [Full
Text] Wagner AD, Maril A, Bjork RA, Schacter DL.
Prefrontal
contributions to executive control: fMRI evidence for functional distinctions
within lateral Prefrontal cortex.
Neuroimage. 2001 Dec;14(6):1337-47.
"The
prefrontal cortex (PFC) plays a fundamental role in internally guided behavior.
Although it is generally accepted that PFC subserves working memory and executive
control operations, it remains unclear whether the subregions within lateral PFC
support distinct executive control processes. An event-related fMRI study was
implemented to test the hypothesis that ventrolateral and dorsolateral PFC are
functionally distinct, as well as to assess whether functional specialization
exists within ventrolateral PFC. Participants performed two executive control
tasks that differed in the types of control processes required. During rote rehearsal,
participants covertly rehearsed three words in the order presented, thus requiring
phonological access and maintenance. During elaborative rehearsal, participants
made semantic comparisons between three words held in working memory, reordering
them from least to most desirable. Thus, in addition to maintenance, elaborative
rehearsal required goal-relevant coding of items in working memory ("monitoring")
and selection from among the items to implement their reordering. Results revealed
that left posterior ventrolateral PFC was active during performance of both tasks,
whereas right dorsolateral PFC was differentially engaged during elaborative rehearsal.
The temporal characteristics of the hemodynamic responses further suggested that
dorsolateral activation lagged ventrolateral activation. Finally, differential
activation patterns were observed within left ventrolateral PFC, distinguishing
between posterior and anterior regions. These data suggest that anatomically separable
subregions within lateral PFC may be functionally distinct and are consistent
with models that posit a hierarchical relationship between dorsolateral and ventrolateral
regions such that the former monitors and selects goal-relevant representations
being maintained by the latter. [Abstract]
[PDF] Miller,
Earl K., Cohen, Jonathan D.
AN INTEGRATIVE THEORY OF PREFRONTAL
CORTEX FUNCTION
Annu. Rev. Neurosci. 2001 24: 167-202
"The prefrontal cortex has long been suspected to play an important role
in cognitive control, in the ability to orchestrate thought and action in accordance
with internal goals. Its neural basis, however, has remained a mystery. Here,
we propose that cognitive control stems from the active maintenance of patterns
of activity in the prefrontal cortex that represent goals and the means to achieve
them. They provide bias signals to other brain structures whose net effect is
to guide the flow of activity along neural pathways that establish the proper
mappings between inputs, internal states, and outputs needed to perform a given
task. We review neurophysiological, neurobiological, neuroimaging, and computational
studies that support this theory and discuss its implications as well as further
issues to be addressed." [Abstract]
[Full Text PDF]
Hunter
MD, Farrow TF, Papadakis NG, Wilkinson ID, Woodruff PW, Spence SA.
Approaching
an ecologically valid functional anatomy of spontaneous "willed" action.
Neuroimage.
2003 Oct;20(2):1264-9.
"We used functional magnetic resonance imaging
of healthy subjects to investigate the neural basis for spontaneous "willed"
action. We hypothesised that such action involves prefrontal cortex (PFC) and
supplementary motor area (SMA), in addition to primary motor cortex. Furthermore,
we predicted that PFC and SMA would demonstrate similar temporal response dynamics,
distinct from primary motor cortex. Specifically, we predicted earlier activation
in PFC and SMA, manifest as shorter response latencies compared with primary motor
cortex. Six right-handed males participated in an event-related design and were
required to generate spontaneous motor acts inside the scanner. By deciding "which"
of two buttons to press, and "when" to press them, subjects generated
sequences of action that were of high information content ("novelty"
or "randomness"). Utilising a short repetition time (1 s), we acquired
functional images that covered most of the frontal and parietal cortices. The
onset of action was associated with significant activation in bilateral PFC, left
primary motor cortex, and, close to the midline, SMA. Following action, mean time
to half-maximum blood oxygenation level-dependent response was significantly earlier
in left PFC and SMA than primary motor cortex. Our findings suggest that neural
correlates of spontaneous willed action are distributed in executive and motor
centres, and that temporal response dynamics differentiate "higher"
regions from subordinate motor areas." [Abstract] St
Clair Gibson A, Baden D, Lambert M, Lambert E, Harley Y, Hampson D, Russell V,
Noakes T.
The Conscious Perception of the Sensation of Fatigue.
Sports
Med 2003;33(3):167-176
"In this review, fatigue is described as a conscious
sensation rather than a physiological occurrence. We suggest that the sensation
of fatigue is the conscious awareness of changes in subconscious homeostatic control
systems, and is derived from a temporal difference between subconscious representations
of these homeostatic control systems in neural networks that are induced by changes
in the level of activity. These mismatches are perceived by consciousness-producing
structures in the brain as the sensation of fatigue. In this model, fatigue is
a complex emotion affected by factors such as motivation and drive, other emotions
such as anger and fear, and memory of prior activity. It is not clear whether
the origin of the conscious sensation of fatigue is associated with particular
localised brain structures, or is the result of electrophysiological synchronisation
of entire brain activity." [Abstract] Moores
E, Laiti L, Chelazzi L.
Associative knowledge controls deployment
of visual selective attention.
Nat Neurosci 2003 Feb;6(2):182-189
"According
to some models of visual selective attention, objects in a scene activate corresponding
neural representations, which compete for perceptual awareness and motor behavior.
During a visual search for a target object, top-down control exerted by working
memory representations of the target's defining properties resolves competition
in favor of the target. These models, however, ignore the existence of associative
links among object representations. Here we show that such associations can strongly
influence deployment of attention in humans. In the context of visual search,
objects associated with the target were both recalled more often and recognized
more accurately than unrelated distractors. Notably, both target and associated
objects competitively weakened recognition of unrelated distractors and slowed
responses to a luminance probe. Moreover, in a speeded search protocol, associated
objects rendered search both slower and less accurate. Finally, the first saccades
after onset of the stimulus array were more often directed toward associated than
control items." [Abstract] Bar,
Moshe
A Cortical Mechanism for Triggering Top-Down Facilitation in
Visual Object Recognition
J. Cogn. Neurosci. 2003 15: 600-609
"The
majority of the research related to visual recognition has so far focused on bottom-up
analysis, where the input is processed in a cascade of cortical regions that analyze
increasingly complex information. Gradually more studies emphasize the role of
top-down facilitation in cortical analysis, but it remains something of a mystery
how such processing would be initiated. After all, top-down facilitation implies
that high-level information is activated earlier than some relevant lower-level
information. Building on previous studies, I propose a specific mechanism for
the activation of top-down facilitation during visual object recognition. The
gist of this hypothesis is that a partially analyzed version of the input image
(i.e., a blurred image) is projected rapidly from early visual areas directly
to the prefrontal cortex (PFC). This coarse representation activates in the PFC
expectations about the most likely interpretations of the input image, which are
then back-projected as an "initial guess" to the temporal cortex to
be integrated with the bottom-up analysis. The top-down process facilitates recognition
by substantially limiting the number of object representations that need to be
considered. Furthermore, such a rapid mechanism may provide critical information
when a quick response is necessary." [Abstract]
[PDF] Sussman
E, Winkler I, Schroger E.
Top-down control over involuntary attention
switching in the auditory modality.
Psychon Bull Rev. 2003
Sep;10(3):630-7.
"We tested the effects of predictability on involuntary
attention switching to task-irrelevant sound changes (distraction). Behavioral
and neurophysiological evidence are provided, showing that the predictability
of task-irrelevant sound changes eliminates effects of distraction even though
the automatic auditory change detection system remains responsive. Two indices
of distraction, slower task performance and cortical brain responses associated
with attention switching, were seen only in the unpredictable condition, in which
the irrelevant acoustic changes were unexpected. Attention was not involuntarily
drawn away from the primary task when the subjects had foreknowledge of when the
irrelevant changes would occur. These results demonstrate attentional control
over orienting to sound changes and suggest that involuntary attention switching
occurs mainly when an irrelevant stimulus change is unexpected. The present data
allowed observation of the temporal dynamics of attention switching in the human
brain." [Abstract]
Liu
T, Slotnick SD, Serences JT, Yantis S.
Cortical mechanisms of feature-based
attentional control.
Cereb Cortex. 2003 Dec; 13(12): 1334-43.
"A
network of fronto-parietal cortical areas is known to be involved in the control
of visual attention, but the representational scope and specific function of these
areas remains unclear. Recent neuroimaging evidence has revealed the existence
of both transient (attention-shift) and sustained (attention-maintenance) mechanisms
of space-based and object-based attentional control. Here we investigate the neural
mechanisms of feature-based attentional control in human cortex using rapid event-related
functional magnetic resonance imaging (fMRI). Subjects viewed an aperture containing
moving dots in which dot color and direction of motion changed once per second.
At any given moment, observers attended to either motion or color. Two of six
motion directions and two of six colors embedded in the stimulus stream cued subjects
either to shift attention from the currently attended to the unattended feature
or to maintain attention on the currently attended feature. Attentional modulation
of the blood oxygenation level dependent (BOLD) fMRI signal was observed in early
visual areas that are selective for motion and color. More importantly, both transient
and sustained BOLD activity patterns were observed in different fronto-parietal
cortical areas during shifts of attention. We suggest these differing temporal
profiles reflect complementary roles in the control of attention to perceptual
features." [Abstract] Ullsperger
M, von Cramon DY, Muller NG.
Interactions of focal cortical lesions
with error processing: evidence from event-related brain potentials.
Neuropsychology.
2002 Oct;16(4):548-61.
"Electrophysiological and hemodynamic studies have
suggested that structures in the vicinity of the anterior cingulate cortex are
involved in performance monitoring, particularly in detection of errors. Bidirectional
interactions between the frontomedian system involved in performance monitoring
and the lateral prefrontal cortex as well as the orbitofrontal cortex have been
proposed, but few studies have directly addressed this issue. The authors used
a speeded flankers task to investigate error-related event-related potentials
in 3 patient groups with different focal cortical lesions. Whereas bilateral frontopolar
lesions involving the orbitofrontal cortex as well as temporal lesions did not
alter the error-related negativity (ERN), lesions of the lateral frontal cortex
resulted in an abolition of the ERN and in a reduction of the error positivity."
[Abstract] Bishop
S, Duncan J, Brett M, Lawrence AD.
Prefrontal cortical function and
anxiety: controlling attention to threat-related stimuli.
Nat
Neurosci. 2004 Feb;7(2):184-8. Epub 2004 Jan 04.
"Threat-related stimuli
are strong competitors for attention, particularly in anxious individuals. We
used functional magnetic resonance imaging (fMRI) with healthy human volunteers
to study how the processing of threat-related distractors is controlled and whether
this alters as anxiety levels increase. Our work builds upon prior analyses of
the cognitive control functions of lateral prefrontal cortex (lateral PFC) and
anterior cingulate cortex (ACC). We found that rostral ACC was strongly activated
by infrequent threat-related distractors, consistent with a role for this area
in responding to unexpected processing conflict caused by salient emotional stimuli.
Participants with higher anxiety levels showed both less rostral ACC activity
overall and reduced recruitment of lateral PFC as expectancy of threat-related
distractors was established. This supports the proposal that anxiety is associated
with reduced top-down control over threat-related distractors. Our results suggest
distinct roles for rostral ACC and lateral PFC in governing the processing of
task-irrelevant, threat-related stimuli, and indicate reduced recruitment of this
circuitry in anxiety." [Abstract] Hasselmo
ME.
Neuromodulation and cortical function: modeling the physiological
basis of behavior.
Behav Brain Res 1995 Feb;67(1):1-27
"Neuromodulators including acetylcholine, norepinephrine, serotonin, dopamine
and a range of peptides alter the processing characteristics of cortical networks
through effects on excitatory and inhibitory synaptic transmission, on the adaptation
of cortical pyramidal cells, on membrane potential, on the rate of synaptic modification,
and on other cortical parameters. Computational models of self-organization and
associative memory function in cortical structures such as the hippocampus, piriform
cortex and neocortex provide a theoretical framework in which the role of these
neuromodulatory effects can be analyzed. Neuromodulators such as acetylcholine
and norepinephrine appear to enhance the influence of synapses from afferent fibers
arising outside the cortex relative to the synapses of intrinsic and association
fibers arising from other cortical pyramidal cells. This provides a continuum
between a predominant influence of external stimulation to a predominant influence
of internal recall (extrinsic vs. intrinsic). Modulatory influence along this
continuum may underlie effects described in terms of learning and memory, signal
to noise ratio, and attention." [Abstract] Polk
TA, Simen P, Lewis RL, Freedman E.
A computational approach to
control in complex cognition.
Brain Res Cogn Brain Res
2002 Dec;15(1):71-83
"Cognitive deficits associated with dorsolateral
prefrontal cortex (DLPFC) damage are often most apparent in higher cognitive tasks
that involve problem solving and managing multiple goals. However, computational
models of prefrontal deficits on such tasks are difficult to construct. Problem
solving is most naturally modeled with symbolic systems (e.g. production systems),
but the effects of lesions are most naturally modeled with subsymbolic systems
(neural networks). We show that when we adopt a simple and plausible model of
neural computation, there is a natural and explicit mapping from symbolic, goal-driven
cognition onto neural computation. We exploit this mapping to construct a neural
network model that is capable of solving complex problems in the Tower of London
task. The model leads to a specific hypothesis about the role of DLPFC in such
tasks, namely, that DLPFC represents internally generated subgoals that modulate
competition among posterior representations. When intact, the model accurately
simulates the behavior of college students even on the most difficult problems.
Furthermore, when the subgoal component is lesioned, it accurately simulates the
behavior of prefrontal patients, including the fact that their deficits are most
apparent on the most difficult tasks and that they have special difficulty with
tasks that require inhibiting a prepotent response." [Abstract] Cameron
S. Carter, Angus M. Macdonald, Matthew Botvinick, Laura L. Ross, V. Andrew Stenger,
Douglas Noll, and Jonathan D. Cohen
Parsing executive processes:
Strategic vs. evaluative functions of the anterior cingulate cortex
PNAS
97: 1944-1948, 2000.
"Under conditions in which their performance indicated
that subjects were engaging strategic processes to reduce the effects of response
conflict, no increased activity was observed in the ACC. This is inconsistent
with the theory that the ACC implements strategic processes to minimize the degree
of conflict elicited by the task. In contrast, when subjects' performance indicated
that strategic processes were less engaged and conflict high, a transient increase
in activity was observed in this region of the brain. This result suggests that
the ACC performs an evaluative function, reflecting the degree of response conflict
elicited by the task. Because the ERN literature suggests that ACC activity is
associated with subsequent "corrective" actions, it is likely that other
components of the neural network implementing executive functions are influenced
by ACC activity to implement strategic processes. This is consistent with the
hypothesized central role for this brain region in the executive control of cognition
(27-29). In the present view, the ACC would serve this function by providing an
on-line conflict signal, indicating the need to engage brain regions such as dorsolateral
prefrontal cortex and IPL to implement strategic processes (2). As such, the ACC
would serve as one component of an "error prevention" network. Future
functional neuroimaging studies are likely to reveal further details of the modular
organization of the neural network subserving executive processes and the mechanisms
by which individual components interact to maintain the tightly coordinated yet
highly flexible activity that characterizes the normal human cognitive system."
[Full Text]
[In light of PCT, this paragraph may be better understood if "response
conflict" is replaced with "conflict between control systems." ACC refers to the anterior cingulate cortex; IPL refers to the inferior parietal
cortex.]
Cabeza R, Nyberg L.
Imaging
cognition II: An empirical review of 275 PET and fMRI studies.
J
Cogn Neurosci 2000 Jan;12(1):1-47
"Positron emission tomography (PET)
and functional magnetic resonance imaging (fMRI) have been extensively used to
explore the functional neuroanatomy of cognitive functions. Here we review 275
PET and fMRI studies of attention (sustained, selective, Stroop, orientation,
divided), perception (object, face, space/motion, smell), imagery (object, space/motion),
language (written/spoken word recognition, spoken/no spoken response), working
memory (verbal/numeric, object, spatial, problem solving), semantic memory retrieval
(categorization, generation), episodic memory encoding (verbal, object, spatial),
episodic memory retrieval (verbal, nonverbal, success, effort, mode, context),
priming (perceptual, conceptual), and procedural memory (conditioning, motor,
and nonmotor skill learning). To identify consistent activation patterns associated
with these cognitive operations, data from 412 contrasts were summarized at the
level of cortical Brodmann's areas, insula, thalamus, medial-temporal lobe (including
hippocampus), basal ganglia, and cerebellum. For perception and imagery, activation
patterns included primary and secondary regions in the dorsal and ventral pathways.
For attention and working memory, activations were usually found in prefrontal
and parietal regions. For language and semantic memory retrieval, typical regions
included left prefrontal and temporal regions. For episodic memory encoding, consistently
activated regions included left prefrontal and medial temporal regions. For episodic
memory retrieval, activation patterns included prefrontal, medial temporal, and
posterior midline regions. For priming, deactivations in prefrontal (conceptual)
or extrastriate (perceptual) regions were consistently seen. For procedural memory,
activations were found in motor as well as in non-motor brain areas. Analysis
of regional activations across cognitive domains suggested that several brain
regions, including the cerebellum, are engaged by a variety of cognitive challenges.
These observations are discussed in relation to functional specialization as well
as functional integration." [Abstract]
Hasegawa, Ryohei P., Blitz, Ari M., Geller, Nancy
L., Goldberg, Michael E.
Neurons in Monkey Prefrontal Cortex That
Track Past or Predict Future Performance
Science 2000 290:
1786-1789
"Although frontal cortex is thought to be important in controlling
behavior across long periods of time, most studies of this area concentrate on
neuronal responses instantaneously relevant to the current task. In order to investigate
the relationship of frontal activity to behavior over longer time periods, we
trained rhesus monkeys on a difficult oculomotor task. Their performance fluctuated
during the day, and the activity of prefrontal neurons, even measured while the
monkeys waited for the targets to appear at the beginning of each set of trials,
correlated with performance in a probabilistic rather than a determinist manner:
neurons reflected past or predicted future performance, much more than they reflected
current performance. We suggest that this activity is related to processes such
as arousal or motivation that set the tone for behavior rather than controlling
it on a millisecond basis, and could result from ascending pathways that utilize
slow, second-messenger synaptic processes.
...
The correlation of neuronal
activity in the prefrontal cortex with general probabilities of behavior with
significant time lags, rather than with the details of the current trial may provide
the cortical neurophysiological function of ascending pathways such as the modulatory
dopaminergic and/or noradrenergic pathways. Usher et al. (13) showed that the
activity and synchrony of neurons in the locus coeruleus correlate with the monkey's
performance. The ascending pathways are clearly important to prefrontal function.
Subcortical aminergic areas project diffusely to prefrontal cortex and receive
input from it (14). A few studies suggest a functional role for them on prefrontal
activity: antagonists change the properties of prefrontal neurons in monkeys performing
memory tasks (15-17); extracellular dopamine increases when a monkey performs
a delayed alternation task, a classic task which has been shown to require the
prefrontal cortex (18). Because these ascending systems work through second-messenger
pathways, these synaptic effects can take place over minutes rather than the few
milliseconds needed for direct short-term synaptic processes and may explain the
time lags seen in our data (19). One can easily postulate that our "past"
neurons receive feedback from the task-related neurons that accomplished the task,
and we can also easily postulate that our "future" neurons feed forward
to neurons that will be responsible for accomplishing the task. If those projections
include a second-messenger step, then we would expect to see the delays that we
have demonstrated. The activity that we have demonstrated
in the prefrontal cortex may set the tone for behavior in a general way, similar
to that accomplished by stimulants, fatigue, enthusiasm, arousal, or other influences
through ascending pathways. The critical result of these experiments is the demonstration
of a tonic signal in prefrontal cortex that changes across minutes rather than
milliseconds, and which predicts or tracks the probability of past or future success
rather than the actual details of the behavior itself." [Full
Text] [Note that perception is controlled; behavior is not controlled. This
article is included to provide an example of gain.] Usher,
Marius, Cohen, Jonathan D., Servan-Schreiber, David, Rajkowski, Janusz, Aston-Jones,
Gary
The Role of Locus Coeruleus in the Regulation of Cognitive
Performance
Science 1999 283: 549-554
"Noradrenergic
locus coeruleus (LC) neurons were recorded in monkeys performing a visual discrimination
task, and a computational model was developed addressing the role of the LC brain
system in cognitive performance. Changes in spontaneous and stimulus-induced patterns
of LC activity correlated closely with fluctuations in behavioral performance.
The model explains these fluctuations in terms of changes in electrotonic coupling
among LC neurons and predicts improved performance during epochs of high coupling
and synchronized LC firing. Cross correlations of simultaneously recorded LC neurons
confirmed this prediction, indicating that electrotonic coupling in LC may play
an important role in attentional modulation and the regulation of goal-directed
versus exploratory behaviors." [Full
Text] [Note that "exploratory behavior" is simply an interpretation
of control.] Bernard J. Baars, James Newman, J.G.
Taylor
Neuronal mechanisms of consciousness:
A Relational Global
Workspace framework.
(Pages 269-278 in S. Hameroff, A. Kaszniak,
J. Laukes, Toward a Science of Consciousness II: The second Tucson discussions
and debates. Cambridge, MA: MIT Press, 1998.)
"This paper explores a remarkable
convergence of ideas and evidence, previously presented in separate places by
its authors. That convergence has now become so persuasive that we believe we
are working within substantially the same broad framework. Taylor’s mathematical
papers on neuronal systems involved in consciousness dovetail well with work by
Newman and Baars on the thalamocortical system, suggesting a brain mechanism much
like the global workspace architecture developed by Baars (see references below).
This architecture is relational, in the sense that it continuously mediates the
interaction of input with memory. While our approaches overlap in a number of
ways, each of us tends to focus on different areas of detail. What is most striking,
and we believe significant, is the extent of consensus, which we believe to be
consistent with other contemporary approaches by Weiskrantz, Gray, Crick and Koch,
Edelman, Gazzaniga, Newell and colleagues, Posner, Baddeley, and a number of others.
We suggest that cognitive neuroscience is moving toward a shared understanding
of consciousness in the brain."
...
"The brain stem-thalamocortical
axis supports the state, but not the detailed contents of consciousness, which
are produced by cortex." [Full
Text] On Site Link: Consciousness
Research John G. Taylor
Constructing
the Relational Mind
PSYCHE, 4(10), June 1998
"The
"relational mind" approach to the inner content of consciousness is
developed in terms of various control structures and processing strategies and
their possible neurobiological identifications in brain sites. This leads naturally
to a division of consciousness into a passive and an active part. A global control
structure for the "single strand" aspect of consciousness is proposed
as the thalamo-nucleus reticularis thalami-cortex coupled system, which is related
to experimental data on the electrical stimulation of awareness. Local control,
in terms of excitatory transfer from pre-processing sites to posterior working
memory regions, is supported by data on subliminal perception timing and disambiguation
of poorly defined percepts. The inner content of consciousness is understood as
arising from the resulting relational features between inputs and stored pre-processing
and episodic memories. Strong analogies are drawn between emergent properties
of the model and suggested properties of "raw feels", supporting the
thesis that working memories are the initial sites for the emergence of phenomenal
awareness, and the frontal lobes for its further adumbration in terms of higher
cognitive processing, including the creation of self." [Full
Text] Cho SB, Baars BJ, Newman J.
A
Neural Global Workspace Model for Conscious Attention.
Neural
Netw 1997 Oct 1;10(7):1195-1206
"Considerable progress is being made in
interdisciplinary efforts to develop a general theory of the neural correlates
of consciousness. Developments of Baars' Global Workspace theory over the past
decade are examples of this progress. Integrating experimental data and models
from cognitive psychology, AI and neuroscience, we present a neurocognitive model
in which consciousness is defined as a global integration and dissemination system
- nested in a large-scale, distributed array of specialized bioprocessors - which
controls the allocation of the processing resources of the central nervous system.
It is posited that this global control is effected via cortical 'gating' of a
strategic thalamic nucleus. The basic circuitry of this neural system is reasonably
well understood, and can be modeled, to a first approximation, employing neural
network principles." [Abstract] Llinas
R, Ribary U, Contreras D, Pedroarena C.
The neuronal basis for consciousness.
Philos Trans R Soc Lond B Biol Sci 1998 Nov 29;353(1377):1841-9
"Attempting
to understand how the brain, as a whole, might be organized seems, for the first
time, to be a serious topic of inquiry. One aspect of its neuronal organization
that seems particularly central to global function is the rich thalamocortical
interconnectivity, and most particularly the reciprocal nature of the thalamocortical
neuronal loop function. Moreover, the interaction between the specific and non-specific
thalamic loops suggests that rather than a gate into the brain, the thalamus represents
a hub from which any site in the cortex can communicate with any other such site
or sites. The goal of this paper is to explore the basic assumption that large-scale,
temporal coincidence of specific and non-specific thalamic activity generates
the functional states that characterize human cognition." [Abstract]
Cudeiro-Mazaira FJ, Rivadulla-Fernandez
JC.
[The thalamus: a dynamic door to perception]
Rev Neurol 2002 Jan 16-31;34(2):121-30
"Far from behaving as a simple
relay station, the thalamic circuits represent the framework on which to build
a truly dynamic biological filter which can select the messages to reach the cortex
according to their relevance and the behavioural state of the person. Such filtering
implies the participation of many neurotransmitters which represent as many different
systems. Outstanding amongst these are the groups formed by acetylcholine and
nitric oxide axons arising from the brainstem, and corticofugal fibers which are
sent back from the cortex to the thalamus." [Abstract] Jones
EG.
Viewpoint: the core and matrix of thalamic organization.
Neuroscience 1998 Jul;85(2):331-45
"The integration of the whole cerebral
cortex and thalamus during forebrain activities that underlie different states
of consciousness, requires pathways for the dispersion of thalamic activity across
many cortical areas. Past theories have relied on the intralaminar nuclei as the
sources of diffuse thalamocortical projections that could facilitate spread of
activity across the cortex. A case is made for the presence of a matrix of superficially-projecting
cells, not confined to the intralaminar nuclei but extending throughout the whole
thalamus. These cells are distinguished by immunoreactivity for the calcium-binding
protein, D28K calbindin, are found in all thalamic nuclei of primates and have
increased numbers in some nuclei. They project to superficial layers of the cerebral
cortex over relatively wide areas, unconstrained by architectonic boundaries.
They generally receive subcortical inputs that lack the topographic order and
physiological precision of the principal sensory pathways. Superimposed upon the
matrix in certain nuclei only, is a core of cells distinguished by immunoreactivity
for another calcium-binding protein, parvalbumin, These project in highly ordered
fashion to middle layers of the cortex in an area-specific manner. They are innervated
by subcortical inputs that are topographically precise and have readily identifiable
physiological properties. The parvalbumin cells form the basis for sensory and
other inputs that are to be used as a basis for perception. The calbindin cells,
especially when recruited by corticothalamic connections, can form a basis for
the engagement of multiple cortical areas and thalamic nuclei that is essential
for the binding of multiple aspects of sensory experience into a single framework
of consciousness." [Abstract] Fernandez
de Molina y Canas A.
[Thalamo-cortical system and consciousness]
An R Acad Nac Med (Madr) 2000;117(4):855-69; discussion 869-81
"After
reviewing the concept of the specific and non specific thalamo-cortical systems,
the connectivity of the relay and intralaminar nuclei is analyzed as well as the
recent data concerning the chemical identity of thalamic neurones, the concept
and distribution of "matrix" and "core" neurones and its functional
role. The intrinsic electrical properties of thalamic neurones, its mode of discharge--depending
of the membrane potential level--and its functional significance in the context
of the brain's global activity are discussed. Of special interest are the studies
on the effects of lesion of the relay and intralaminar nuclei as well as its repercussion
in the interpretation of the sensory perception. After intralaminar nuclei lesion
the individual is not aware of the information conveyed through the specific channels.
It follows a discussion on the importance of the temporal and spatial mapping
in the elaboration of perception and cognition. Due to the intrinsic electrical
properties and the connectivity of thalamic neurones two groups of corticothalamic
loops are generated, which resonate at a frequency of 40 Hz. The specific thalamo-cortical
loops give the content of cognition and the non specific loop, the temporal binding
required for the unity of the cognitive experience. Consciousness is then, a product
of the resonant thalamo-cortical activity, and the dialogue between the thalamus
and cortex, the process that generates subjectivity, the unique experience we
all recognized as the existence of the "self"." [Abstract] Balkin
TJ, Braun AR, Wesensten NJ, Jeffries K, Varga M, Baldwin P, Belenky G, Herscovitch
P.
The process of awakening: a PET study of regional brain activity
patterns mediating the re-establishment of alertness and consciousness.
Brain 2002 Oct;125(Pt 10):2308-19
"Awakening from sleep entails rapid
re-establishment of consciousness followed by the relatively slow (20-30 min later)
re-establishment of alertness--a temporal dissociation that facilitates specification
of the physiological underpinnings of each of these facets of the awakening process.
H(2)(15)O PET was used to assess changes in regional cerebral blood flow (rCBF)
upon awakening from stage 2 sleep. Cerebral blood flow (CBF) was most rapidly
re-established in centrencephalic regions (e.g. brainstem and thalamus), suggesting
that the reactivation of these regions underlies the re-establishment of conscious
awareness. Across the ensuing 15 min of wakefulness, further increases in CBF
were evident primarily in anterior cortical regions, suggesting that the dissipation
of sleep inertia effects (post-awakening performance and alertness deficits) is
effected by reactivation of these regions. Concomitant shifts in correlation patterns
of regional brain activity across the post-awakening period [in particular, a
waning negative correlation between prefrontal cortex and mesencephalic reticular
formation (RF) activity, and a waxing positive correlation between prefrontal
cortex and ventromedial caudate nucleus (CAUD) activity] suggest that the post-awakening
reversal of sleep inertia effects may be mediated by more than mere reactivation--it
may also involve the functional reorganization of brain activity. Conversely,
stable post-awakening correlations--such as those found between the anterior cingulate
cortex (ACC) and most other brain regions--may denote the pattern of functional
connectivity that underlies consciousness itself." [Abstract] Ergenzinger
ER, Glasier MM, Hahm JO, Pons TP.
Cortically induced thalamic plasticity in the primate somatosensory
system.
Nat Neurosci 1998 Jul;1(3):226-9
"The
influence of cortical feedback on receptive field organization in the thalamus
was assessed in the primate somatosensory system. Chronic and acute suppression
of neuronal activity in primary somatosensory cortex resulted in a striking enlargement
of receptive fields in the ventroposterior thalamus. This finding demonstrates
a dramatic 'top-down' influence of cortex on receptive field size in the somatosensory
thalamus. In addition, this result has important implications for studies of adult
neuronal plasticity because it indicates that changes in 'higher-order' areas
of the brain can trigger extensive changes in the receptive field characteristics
of neurons located earlier in the processing pathway." [Abstract] Guillery
RW.
Anatomical evidence concerning the role of the thalamus in corticocortical
communication: a brief review.
J Anat 1995 Dec;187 ( Pt
3):583-92
"Two distinct types of thalamic nucleus are proposed on the
basis of the afferent fibres that they receive from ascending pathways and from
the cerebral cortex. 'First order nuclei' receive primary afferent fibres, definable
on the basis of their origin and their intrathalamic synaptic relationships, from
ascending pathways. These nuclei receive corticothalamic afferents from pyramidal
cells in cortical layer 6, which also send branches to the thalamic reticular
nucleus and appear to have a modulatory function. 'Higher order nuclei' receive
most or all of their 'primary afferents' from pyramidal cells in cortical layer
5. These resemble the ascending primary afferents in the first order nuclei in
terms of fine structure, synaptic relationships and in lacking a branch to the
thalamic reticular nucleus. The higher order nuclei also receive modulatory afferents
from layer 6. It is proposed that the higher-order nuclei are largely concerned
with transmitting information about the output of one cortical area to another
cortical area, and that they are likely to play a key role in corticocortical
communication and higher cortical functions." [Abstract] McAlonan
K, Brown VJ.
The thalamic reticular nucleus: more than a sensory
nucleus?
Neuroscientist 2002 Aug;8(4):302-5
"Sensory
information is routed to the cortex via the thalamus, but despite this sensory
bombardment, animals must attend selectively to stimuli that signal danger or
opportunity. Sensory input must be filtered, allowing only behaviorally relevant
information to capture limited attentional resources. Located between the thalamus
and cortex is a thin lamina of neurons called the thalamic reticular nucleus (Rt).
The thalamic reticular nucleus projects exclusively to thalamus, thus forming
an essential component of the circuitry mediating sensory transmission. This article
presents evidence supporting a role for Rt beyond the mere relay of sensory information.
Rather than operating as a component of the sensory relay, the authors suggest
that Rt represents an inhibitory interface or "attentional gate," which
regulates the flow of information between the thalamus and cortex. Recent findings
have also implicated Rt in higher cognitive functions, including learning, memory,
and spatial cognition. Drawing from recent insights into the dynamic nature of
the thalamic relay in awake, behaving animals, the authors present a speculative
account of how Rt might regulate thalamocortical transmission and ultimately the
contents of consciousness." [Abstract]
Castro-Alamancos, Manuel A.
Role
of Thalamocortical Sensory Suppression during Arousal: Focusing Sensory Inputs
in Neocortex
J. Neurosci. 2002 22: 9651-9655
"The
thalamus serves as a gate that regulates the flow of sensory inputs to the neocortex,
and this gate is controlled by neuromodulators from the brainstem reticular formation
that are released during arousal. We found recently that sensory-evoked responses
are suppressed in the neocortex during arousal. This sensory suppression results
from the activity-dependent depression of the thalamocortical connection caused
by increased tonic firing of thalamocortical cells during arousal. In the present
study, the functional consequences of thalamocortical suppression during arousal
were investigated using the vibrissae system of rodents. The results show that
thalamocortical suppression is associated with a strong reduction in the spread
of sensory inputs through the cortex, thus reducing the size of sensory representations.
In addition, when the responses of single cells to principal and adjacent whiskers
are compared, the response to the adjacent whiskers was found to be strongly suppressed,
much more so than that of principal whiskers. Consequently, the receptive fields
of cortical neurons become more focused to the principal whisker. The results
indicate that thalamocortical suppression during arousal serves to focus sensory
inputs to their appropriate representations in neocortex, which may be computationally
helpful for the spatial processing of sensory information." [Abstract]
Smythies J.
The functional neuroanatomy
of awareness: with a focus on the role of various anatomical systems in the control
of intermodal attention.
Conscious Cogn 1997 Dec;6(4):455-81
"This review considers a number of recent theories on the neural basis of
consciousness, with particular attention to the theories of Bogen, Crick, Llinas,
Newman, and Changeux. These theories allot different roles to various key brain
areas, in particular the reticular and intralaminar nuclei of the thalamus and
the cortex. Crick's hypothesis is that awareness is a function of reverberating
corticothalamic loops and that the spotlight of intramodal attention is controlled
by the reticular nucleus of the thalamus. He also proposed different mechanisms
for attention and intention ("will"). The current review presents a
new hypothesis, based on elements from these hypotheses, including intermodal
attention and olfaction and pain, which may pose problems for Crick's original
theory. This work reviews the possible role in awareness and intermodal attention
and intention of the cholinergic system in the basal forebrain and the tegmentum;
the reticular, the intralaminar, and the dorsomedial thalamic nuclei; the raphe
and locus coeruleus; the reticular formation; the ventral striatum and extended
amygdala; insula cortex, and other selected cortical, areas. Both clinical and
basic research data are covered. The conclusion is reached that the brain may
work by largely nonlinear parallel processing and much intramodal shifts of attention
may be effected by intracortical, or multiple corticothalamic mechanisms (small
local "flashlights" rather than one major "searchlight").
But this is constrained by the functional anatomy of the circuits concerned and
waking "awareness" is modulated by the many "nonspecific"
systems (cholinergic from the basal forebrain, noradrenergic from the locus coeruleus,
dopaminergic from the substantia nigra and ventral tegmentum, and serotoninergic
from the raphe). But the principal agents for intermodal attention shifts, the
"searchlight," may be two key nuclei of the cholinergic system in the
mesencephalon. Clinical loss of consciousness results from damage to these nuclei
but not from damage to the cholinergic nucleus basalis of the basal forebrain."
[Abstract] Perry
E, Walker M, Grace J, Perry R.
Acetylcholine in mind: a neurotransmitter
correlate of consciousness?
Trends Neurosci 1999 Jun;22(6):273-80
"The cholinergic system is one of the most important modulatory neurotransmitter
systems in the brain and controls activities that depend on selective attention,
which are an essential component of conscious awareness. Psychopharmacological
and pathological evidence supports the concept of a 'cholinergic component' of
conscious awareness. Drugs that antagonize muscarinic receptors induce hallucinations
and reduce the level of consciousness, while the nicotinic receptor is implicated
as being involved in the mechanism of action of general (inhalational) anaesthetics.
In degenerative diseases of the brain, alterations in consciousness are associated
with regional deficits in the cholinergic system. In Alzheimer's disease (AD),
there is a loss of explicit (more than implicit) memory and hypoactivity of cholinergic
projections to the hippocampus and cortex, while the visual hallucinations experienced
by subjects with Dementia with Lewy bodies (DLB) are associated with reductions
in neocortical ACh-related activity. In Parkinson's disease, the additional loss
of pedunculopontine cholinergic neurones, which control REM (rapid eye movement)
sleep or dreaming, is likely to contribute to REM abnormalities, which also occur
in DLB. Widespread basal-forebrain and rostral brainstem cholinergic pathways,
which include converging projections to the thalamus, appear to be located strategically
for generating and integrating conscious awareness. Alleviation of a range of
cognitive and non-cognitive symptoms by drugs that modulate the cholinergic system,
which are being developed for the treatment of AD and related disorders, could
be caused by changes in consciousness." [Abstract] Manuel
A. Castro-Alamancos, and Maria E. Calcagnotto
High-Pass Filtering
of Corticothalamic Activity by Neuromodulators Released in the Thalamus During
Arousal: In Vitro and In Vivo
J Neurophysiol 85: 1489-1497,
2001.
"The thalamus is the principal relay station of sensory information
to the neocortex. In return, the neocortex sends a massive feedback projection
back to the thalamus. The thalamus also receives neuromodulatory inputs from the
brain stem reticular formation, which is vigorously activated during arousal.
We investigated the effects of two neuromodulators, acetylcholine and norepinephrine,
on corticothalamic responses in vitro and in vivo. Results from rodent slices
in vitro showed that acetylcholine and norepinephrine depress the efficacy of
corticothalamic synapses while enhancing their frequency-dependent facilitation.
This produces a stronger depression of low-frequency responses than of high-frequency
responses. The effects of acetylcholine and norepinephrine were mimicked by muscarinic
and alpha(2)-adrenergic receptor agonists and blocked by muscarinic and alpha-adrenergic
antagonists, respectively. Stimulation of the brain stem reticular formation in
vivo also strongly depressed corticothalamic responses. The suppression was very
strong for low-frequency responses, which do not produce synaptic facilitation,
but absent for high-frequency corticothalamic responses. As in vitro, application
of muscarinic and alpha-adrenergic antagonists into the thalamus in vivo abolished
the suppression of corticothalamic responses induced by stimulating the reticular
formation. In conclusion, cholinergic and noradrenergic activation during arousal
high-pass filters corticothalamic activity. Thus, during arousal only high-frequency
inputs from the neocortex are allowed to reach the thalamus. Neuromodulators acting
on corticothalamic synapses gate the flow of cortical activity to the thalamus
as dictated by behavioral state." [Full
Text]
Sturm W, Willmes K.
On
the functional neuroanatomy of intrinsic and phasic alertness.
Neuroimage 2001 Jul;14(1 Pt 2):S76-84
"Intrinsic and phasic alertness
are the most basic aspects of attention intensity probably constituting the basis
for the more complex and capacity-demanding aspects of attention selectivity.
Intrinsic alertness represents the cognitive control of wakefulness and arousal
and is typically assessed by simple reaction time tasks without a preceding warning
stimulus. Phasic alertness, in contrast, is called for in reaction time tasks
in which a warning stimulus precedes the target, and it represents the ability
to increase response readiness subsequent to external cueing. We report PET and
fMRI data from both the literature and our own experiments to delineate the cortical
and subcortical networks subserving alertness, sustained attention (as another
aspect of attention intensity), and spatial orienting of attention. Irrespective
of stimulus modality, there seems to exist a mostly right-hemispheric frontal,
parietal, thalamic, and brain-stem network which is coactivated by alerting and
orienting attentional demands. These findings corroborate both the hypothesis
of a frontal modulation of brain-stem activation probably via the reticular nucleus
of the thalamus and of a coactivation of the posterior attention system involved
in spatial orienting by the anterior alerting network. Under conditions of phasic
alertness there are additional activations of left-hemisphere frontal and parietal
structures which are interpreted as basal aspects of attention selectivity rather
than additional features of alerting." [Abstract] Wolkenhauer
O.
Mathematical modelling in the post-genome era: understanding
genome expression and regulation--a system theoretic approach.
Biosystems 2002 Feb;65(1):1-18
"This paper introduces a mathematical
framework for modelling genome expression and regulation. Starting with a philosophical
foundation, causation is identified as the principle of explanation of change
in the realm of matter. Causation is, therefore, a relationship, not between components,
but between changes of states of a system. We subsequently view genome expression
(formerly known as 'gene expression') as a dynamic process and model aspects of
it as dynamic systems using methodologies developed within the areas of systems
and control theory. We begin with the possibly most abstract but general formulation
in the setting of category theory. The class of models realised are state-space
models, input--output models, autoregressive models or automata. We find that
a number of proposed 'gene network' models are, therefore, included in the framework
presented here. The conceptual framework that integrates all of these models defines
a dynamic system as a family of expression profiles. It becomes apparent that
the concept of a 'gene' is less appropriate when considering mathematical models
of genome expression and regulation. The main claim of this paper is that we should
treat (model) the organisation and regulation of genetic pathways as what they
are: dynamic systems. Microarray technology allows us to generate large sets of
time series data and is, therefore, discussed with regard to its use in mathematical
modelling of gene expression and regulation." [Abstract] Gierer
A.
Networks of gene regulation, neural development and the evolution
of general capabilities, such as human empathy.
Z Naturforsch
[C] 1998 Jul-Aug;53(7-8):716-22
"A network of gene regulation organized
in a hierarchical and combinatorial manner is crucially involved in the development
of the neural network, and has to be considered one of the main substrates of
genetic change in its evolution. Though qualitative features may emerge by way
of the accumulation of rather unspecific quantitative changes, it is reasonable
to assume that at least in some cases specific combinations of regulatory parts
of the genome initiated new directions of evolution, leading to novel capabilities
of the brain. These notions are applied, in this paper, to the evolution of the
capability of cognition-based human empathy. It is suggested that it has evolved
as a secondary effect of the evolution of strategic thought. Development of strategies
depends on abstract representations of one's own possible future states in one's
own brain to allow assessment of their emotional desirability, but also on the
representation and emotional evaluation of possible states of others, allowing
anticipation of their behaviour. This is best achieved if representations of others
are connected to one's own emotional centres in a manner similar to self-representations.
For this reason, the evolution of the human brain is assumed to have established
representations with such linkages. No group selection is involved, because the
quality of strategic thought affects the fitness of the individual. A secondary
effect of this linkage is that both the actual states and the future perspectives
of others elicit vicarious emotions, which may contribute to the motivations of
altruistic behaviour." [Abstract] Lau
HC, Rogers RD, Haggard P, Passingham RE.
Attention to intention.
Science.
2004 Feb 20;303(5661):1208-10.
"Intention is central to the concept of
voluntary action. Using functional magnetic resonance imaging, we compared conditions
in which participants made self-paced actions and attended either to their intention
to move or to the actual movement. When they attended to their intention rather
than their movement, there was an enhancement of activity in the pre-supplementary
motor area (pre-SMA). We also found activations in the right dorsal prefrontal
cortex and left intraparietal cortex. Prefrontal activity, but not parietal activity,
was more strongly coupled with activity in the pre-SMA. We conclude that activity
in the pre-SMA reflects the representation of intention." [Abstract] Rowe
J, Friston K, Frackowiak R, Passingham R.
Attention to action: specific
modulation of corticocortical interactions in humans.
Neuroimage.
2002 Oct;17(2):988-98.
"The prefrontal cortex may exert cognitive control
by a general mechanism of attentional selection of neuronal representations. We
used functional magnetic resonance imaging to test this hypothesis in the motor
system. Normal volunteers were scanned during performance of a simple motor task,
with their attention either directed towards their actions, or diverted towards
a visual search task, or neither. Attention to action increased activity in prefrontal,
premotor and parietal cortex, compared with unattended performance of the same
movements. Analysis of cortical activity by structural equation modelling of regional
fMRI time series was used to measure effective connectivity among prefrontal,
premotor and parietal cortices. Attention to action enhanced effective connectivity
between dorsal prefrontal cortex and premotor cortex, whereas non-motor attention
diminished it. These effects were not attributable to common inputs from parietal
cortex to the prefrontal and premotor cortex. The results suggest a supra-modal
role for the dorsal prefrontal cortex in attentional selection, operating within
the motor system as well as sensory and mnemonic domains." [Abstract]
Jimura
K, Konishi S, Miyashita Y.
Dissociable concurrent activity of lateral
and medial frontal lobe during negative feedback processing.
Neuroimage.
2004 Aug;22(4):1578-86.
External feedback on results of one's behavior guides
flexible adaptation to changing environments. It has been suggested that the lateral
and medial parts of the frontal lobe are responsible for cognitive and emotional
functions, respectively. In the present fMRI study, multiple mental components
evoked by the presentation of negative feedback were dissociated along the cognitive-emotional
axis in set-shifting paradigms. The double dissociation of the concurrent feedback-related
activity was observed in the right frontal lobe: the lateral frontal lobe was
associated with the inferential component, whereas the medial frontal lobe was
associated with the emotional component. However, among the multiple right lateral
frontal regions, a region of interest (ROI) analysis indicated that the inferential
component was not dominant in the region near the inferior frontal junction. The
medial frontal activations were observed ventral and anterior to the presupplementary
motor area, and dorsal and posterior to the anterior cingulate cortex. The double
dissociation in the right frontal lobe suggests that the lateral and medial frontal
lobe cooperatively but differentially contributes to the negative feedback processing,
demonstrating the lateral-medial dichotomy of the frontal lobe functions suggested
by previous neuropsychological studies. At the same time, the functional heterogeneity
in the lateral and medial frontal lobe demands modifications of the traditional
view of the functional organization of the frontal lobe. [Abstract] |
